돼지의 병원체인 Streptococcus suis균은 그람 양성 세균으로 돼지나 돈육제품에 노출된 인간에게 심각한 질병을 일으키는 인수공통전염병 세균입니다. 지난 2005년 중국에서 Streptococcus suis균에 206명이 감염되어 38명이 사망하는 사건이 발생하기도 했는데요… 당시 사망자들(주로 농부들)은 피부 안쪽으로 출혈이 일어나는 괴질의 원인체이기도 했습니다.
이탈리아 연구팀은 PDR기법을 이용하여 2명의 사망자에서 분리된 Streptococcus suis균과 다른 57명의 환자에게서 분리된 Streptococcus suis균을 분석했습니다.
분리된 Streptococcus suis균의 항생제 내성율을 보면 매크로라이드(macrolides) 계열의 항생제 내성이 78%에 달했고, 테트라사이클린 계열의 항생제 내성이 90%에 이르는 것으로 나타납니다.
그리고 Streptococcus suis균의 erm(B), tet(O), mosaic tet(O/W/32/O), tet(W), tet(M) 유전자가 검출되었는데요… tet(O) 다음으로 많이 검출된 tet(O/W/32/O) 유전자는 Streptococcus속( genus)에서는 이번에 처음 발견되었습니다.
[참고] 매크로라이드(macrolides) 계열의 항생제
Erythromycin,roxithrimycin, dirithromycin, clarithromycin, flurithromycin, azithromycin, rokitamycin, leucomycin A5, josamycin, miocamycin, triacetylspiramycin, spiramycin
[참고] 우리나라에서 1998년도 인간에게서 분리된 A군 연쇄구균에 대한 erythromycin 내성균이 46%정도이고 erythromycin에 내성을 갖는 A군 연쇄구균은 서로 교차 내성이 있기 때문에 다른 새로운 macrolides에도 내성이 있는 것으로 알려져 있습니다.
======================================
Research articles
Genetic diversity of Streptococcus suis clinical isolates from pigs and humans in Italy (2003-2007)
출처 : Eurosurveillance, Volume 14, Issue 33, 20 August 2009
http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19310
M S Princivalli1, C Palmieri1, G Magi1, C Vignaroli1, A Manzin2, A Camporese3, S Barocci4, C Magistrali4, B Facinelli (
- Department of Biomedical Sciences, Polytechnic University of Marche Medical School, Ancona, Italy
- Department of Biomedical Sciences and Technologies, Section of Medical Microbiology, University of Cagliari Medical School, Italy
- Microbiology and Virology Department, S. Maria degli Angeli Regional Hospital, Pordenone, Italy
- Experimental Zooprophylactic Institute of Umbria and Marche, Perugia, Italy
Date of submission: 22 April 2009
Streptococcus suis, a major porcine pathogen, is emerging as a zoonotic agent capable of causing severe invasive disease in humans exposed to pigs or pork products. S. suis infection is rare in industrialised countries and usually arises as sporadic cases, with meningitis the most common clinical presentation in humans. Recent reports of two cases of meningitis in Sardinia and north-eastern Italy prompted this first characterisation of Italian S. suis isolates. Fifty-nine S. suis strains, the two recent human strains and 57 swine clinical isolates collected between 2003 and 2007 from different Italian herds and regions, were tested for antimicrobial susceptibility, PCR-screened for virulence and antibiotic resistance genes, and subjected to molecular typing. Phenotypic and genotypic analysis demonstrated an overall high genetic diversity among isolates, the majority of which were resistant to macrolides (78%) and tetracyclines (90%). The erm(B), tet(O), mosaic tet(O/W/32/O), tet(W), and tet(M) genes were detected. The tet(O/W/32/O) gene, the most frequent tet gene after tet(O), had never been described in the genus Streptococcus before. In addition, a virulent cps2, erm(B) tet(O) clone, belonging to sequence type 1 (ST1) of the ST1 complex, was found to be prevalent and persistent in Italian swine herds. Finally, the two human isolates (both ST1) carrying cps2, erm(B) and tet(W) were seen to be closely related to each other.
Introduction
Streptococcus suis, a major porcine pathogen endemic in nearly all countries with a developed swine industry, causes meningitis, pneumonia, arthritis, endocarditis, and septicaemia in pigs [1]. S. suis is also emerging as a zoonotic agent capable of causing severe invasive disease in humans exposed to pigs or to pork products [2,3]. A carriage state has been documented in pigs, healthy carriers being a source of S. suis transmission in herds, mainly through the respiratory route [1]. As discussed in recent reports, the possibility cannot be excluded that humans may also be healthy carriers [1,3,4] and that S. suis may become an opportunistic pathogen under particular circumstances such as stress, immunodeficiency or cancer [1,5]. Meningitis with possible residual deafness is the most frequent clinical presentation of the infection in humans; septicaemia, pneumonia, endocarditis, arthritis and toxic shock syndrome have also been described. In industrialised countries, S. suis disease is rare, albeit probably underdiagnosed, and usually occurs as sporadic cases [2,3]. Most human cases reported so far originated from Southeast Asia, where the disease can be considered endemic and where some outbreaks have occurred [3]. Three major sequence type (ST) clonal complexes (ST1, ST27 and ST87) dominate the population [6]. The virulent ST1 complex, frequently associated with invasive infections, includes sequence type ST1, spread worldwide and recently detected for the first time in Italy [5], and ST7, responsible for several cases of toxic shock syndrome during a recent outbreak in China [7].
The antiphagocytic polysaccharide capsule (encoded by the cps gene) is the major virulence factor of S. suis. Thirty-three serotypes based on capsular antigens are currently recognised [8,9]. Serotype 2 is responsible for severe infections in swine [1] and is the most common serotype affecting humans worldwide [2]. The small number of human S. suis infections in North America has been linked to the low prevalence of serotype 2 among swine [1]. Serotypes 4, 14 and 16 have also been described in humans [1]. Proposed S. suis virulence factors [1], the significance of which is still unknown, include the muramidase released protein MRP (encoded by mrp), a peptidoglycan-associated protein probably acting as an adhesin and the extracellular protein factor EF (epf), both of which are suitable virulence markers of serotype 2 strains [10] and are also detected in other serotypes [11], a serum opacity factor OFS (ofs), proposed as a virulence trait of cps2 isolates [12,13], suilysin (sly), a haemolysin with a cytotoxic effect on various cell types [1], and arginine deiminase (arcA), a factor linked to survival in stress conditions [14]. Despite the lack of evidence for a critical role of one or more of these putative virulence factors in virulence, they may nonetheless serve as virulence markers, since MRP, EF, and suilysin are typical of Eurasian strains of the ST1 complex, while they are almost absent in less virulent North American strains [1]. An immune evasion strategy has recently been proposed to account for the allelic variability observed in mrp, epf, and ofs genes [11,13].
A trend toward mounting S. suis resistance to macrolides and tetracyclines has been reported worldwide [15-17]. Studies of genetic resistance traits have demonstrated erm(B) (ribosomal methylation) and mef(A) (active efflux) for macrolide resistance, and tet(M) and tet(O) (both ribosomal protection) for tetracycline resistance [18-21]. The tet(W) gene, an emerging determinant commonly found in species inhabiting human and animal intestinal tracts [22], was first detected by our group in a human isolate of S. suis from a case of meningitis in Italy [5].
Overall, three human cases of S. suis meningitis have been reported in Italy, one in the 1990s [23] and two quite recently, in the course of little more than a year. The short interval between the last two cases and their arising in distant geographic areas, i.e. north-eastern Italy [24] and Sardinia [5], prompted this first characterisation of Italian S. suis isolates.
Methods
S. suis strains
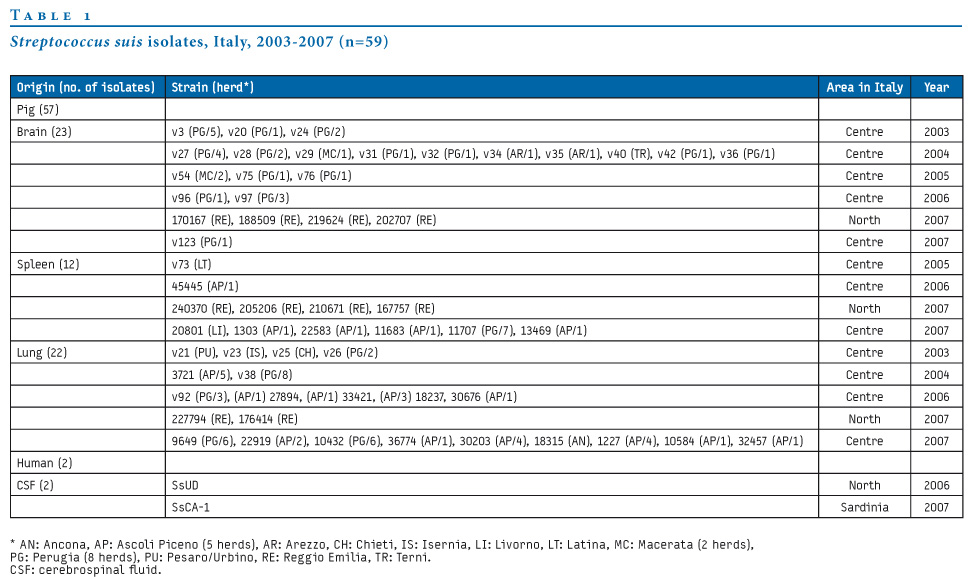
A total of 59 S. suis isolates were studied, two of human and 57 of porcine origin (Table 1). The human isolates, one from Sardinia (SsCA-1: cps2 ST1 erm(B) tet(W)) [5] and the other from north-east Italy [24], here designated as SsUD, were from cerebrospinal fluid (CSF) of two patients with S. suis meningitis. All pig isolates were from clinical samples (23 brain, 22 lung and 12 spleen samples) collected in 24 herds in northern and central Italy from 2003 to 2007. They were divided into invasive (brain and spleen isolates: 35 strains) and non-invasive (lung isolates: 22 strains) according to the source of isolation. All strains were isolated on 5% sheep blood agar (Oxoid Ltd) and identified with ID 32 STREP kit (bioMérieux). Serotyping was performed by slide agglutination using specific antisera (Statens Serum Institute).
Table 1. Streptococcus suis isolates, Italy, 2003-2007 (n=59)
Susceptibility testing
Antimicrobial susceptibility testing by agar disk diffusion and minimal inhibitory concentration (MIC) was carried out according to standard procedures [25,26] (erythromycin and tetracycline antibiotics: Sigma Chemical Co, disks: Oxoid). S. pneumoniae ATCC 49619 was used for quality control. The erythromycin resistance phenotype was determined on the basis of the triple disk test (erythromycin plus clindamycin and josamycin) [27].
Genotyping
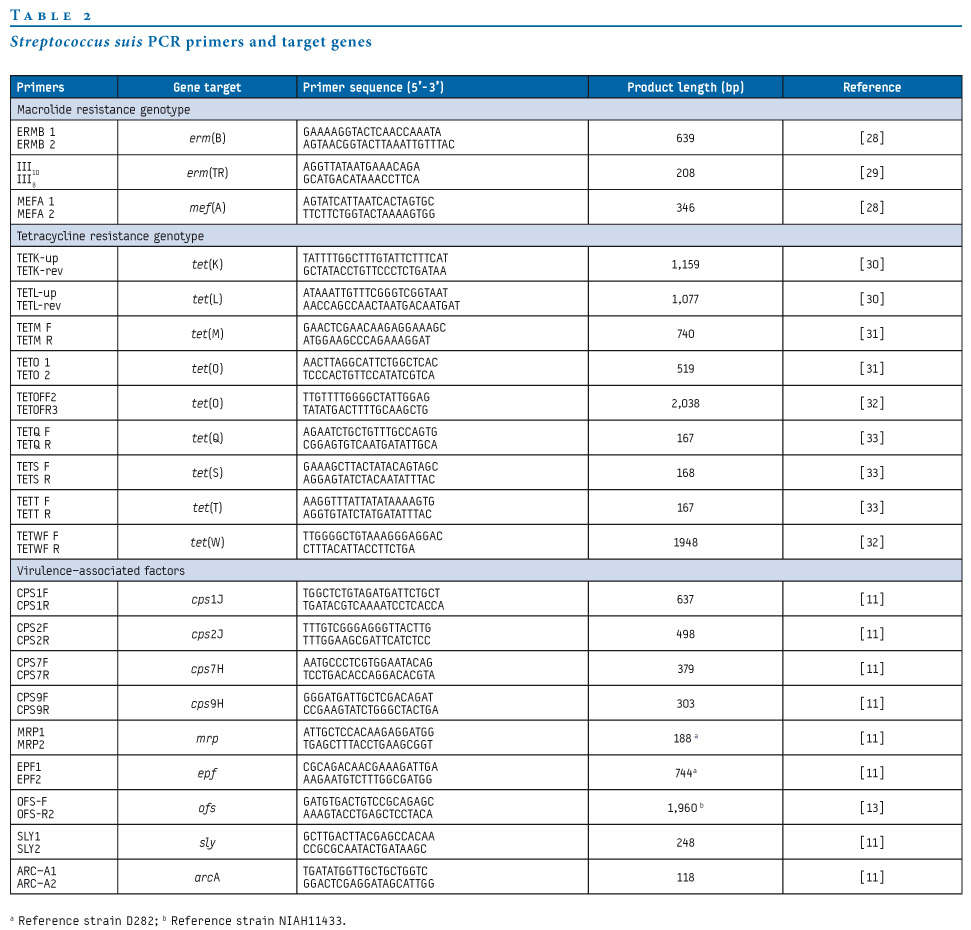
PCR amplification was carried out under published conditions using the oligonucleotide primer pairs and target genes listed in Table 2 [28-33].
Table 2. Streptococcus suis PCR primers and target genes
Pulsed-Field Gel Electrophoresis (PFGE) was applied to study the genetic diversity of S. suis [19,34-36]. Macrorestriction with SmaI endonuclease (Roche) and PFGE analysis were performed essentially as described previously [35]. PFGE data were analysed considering each band as a separate putative locus and scoring it as present (1) or absent (0) in each accession. The dendrogram was constructed by use of the Dice coefficient and the unweighted pair group method with arithmetic averages. Genetic relatedness was interpreted according to the criteria of Tenover et al. [37].
A multilocus sequence typing (MLST) scheme for S. suis was developed in 2002 [6]. Primers for PCR amplification and sequencing of the housekeeping gene fragments of aroA (EPSP synthase), cpn60 (60-kDa chaperonin), dpr (peroxide resistance), gki (glucose kinase), mutS (DNA mismatch repair enzyme), recA (homologous recombination) and thrA (aspartokinase) were synthesised according to the primer sequences on the S. suis MLST database website (http://ssuis.mlst.net). Sequences were compared with previously observed allelic sequences in the S. suis MLST database for identification of ST.
The nucleotide sequences reported here have been submitted to the GenBank/EMBL sequence database and assigned accession numbers FM201280 (ofstype 1S), FN357200 (epf915), FN356743 (tet(W)) and FM164392 (tet(O/W/32/O)). Sequence similarity searches were carried out using BLAST, available online from the National Center for Biotechnology Information of the National Library of Medicine (http://www.ncbi.nlm.nih.gov).
Results
Capsular (cps) and virulence-associated genes
The 59 S. suis isolates were investigated by PCR using primer pairs specific for cps1, cps2, cps7, and cps9, and for virulence-associated genes mrp, epf, ofs, sly, and arcA. Size variants were detected by restriction analysis (epf: HindIII; ofs: MboI) and sequencing (ofs) of PCR products (Table 3). The distributions of cps and virulence-associated genes are reported in the Figure, and virulence profiles among invasive and non-invasive isolates are shown in Table 4.
Table 3. The mrp, epf, and ofs gene size variants observed in Streptococcus suis isolates, Italy, 2003-2007
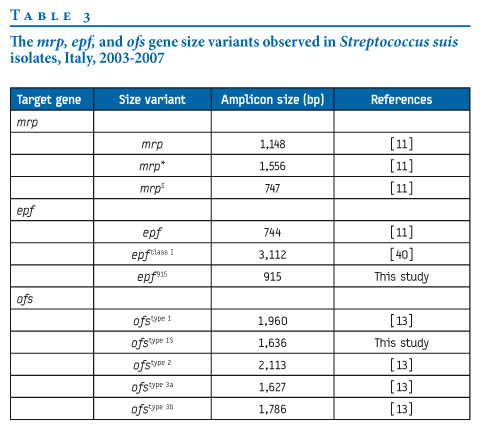
Figure. Similarity index of the 59 Streptococcus suis isolates, Italy, 2003-2007
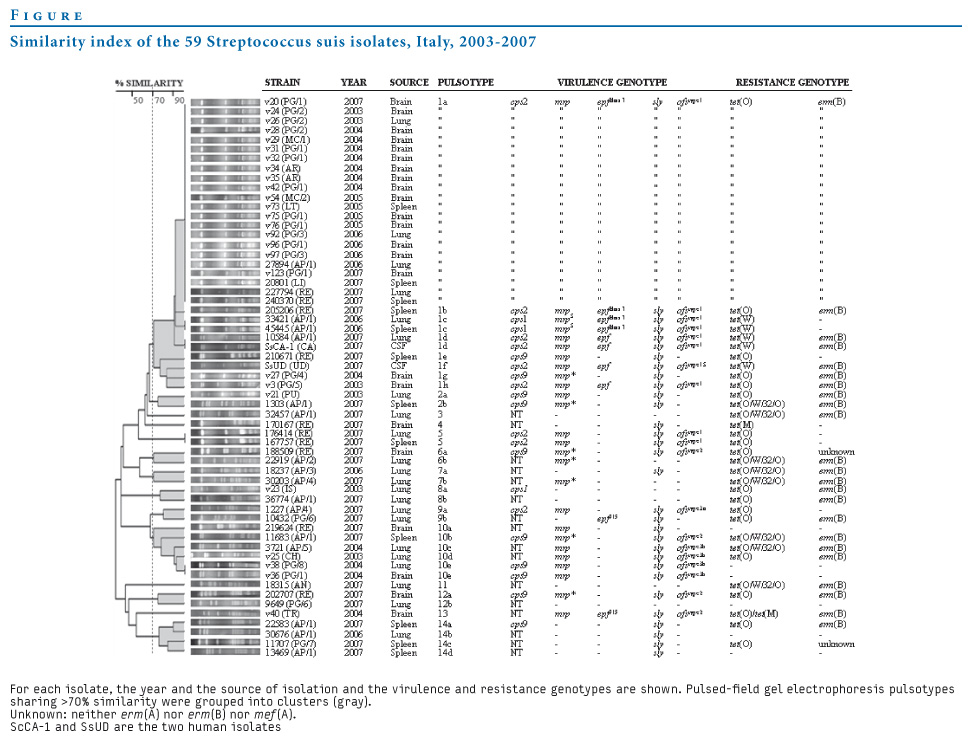
Table 4. Virulence-associated gene profiles in Streptococcus suis isolates, Italy, 2003-2007 (n=59)
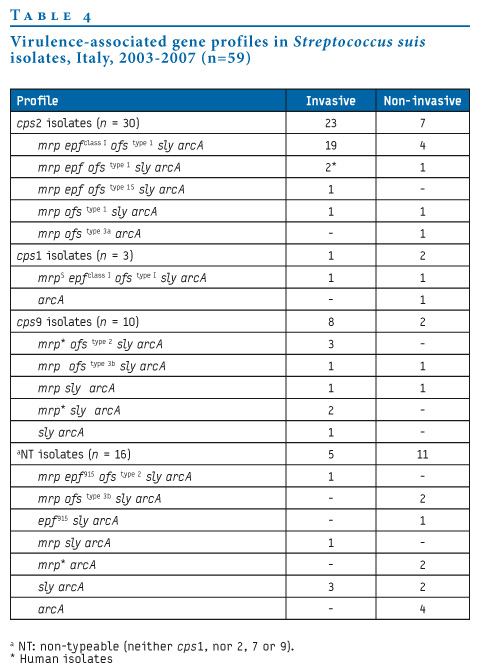
Three cps genes were detected in 43 of the 59 isolates: cps1 (n=3 isolates, one invasive), cps2 (n=30, 23 invasive, including the two human CSF isolates) and cps9 (n=10, eight invasive). In agglutination tests, all cps2 strains showed agglutination with sera specific for serotype 2. The remaining 16 isolates, of which five were invasive, were negative and are referred to as non-typeable (NT).
The mrp gene (three size variants: mrp; mrp* and mrpS) was detected in 47 strains (all 30 cps2 isolates, nine cps9, six NT, and two cps1 isolates); epf (three size variants: epf; epfclass I and epf915) was detected in 31 strains (27 cps2, two cps1 and two NT isolates); ofs (five size variants: ofstype 1, ofstype 1S, ofstype 2, ofstype 3a and ofstype 3b) was detected in 40 strains (all 30 cps2, five cps9, three NT and two cps1 isolates); sly was detected in 52 strains (all cps2 and cps9 isolates, two cps1 and 10 NT isolates), and arcA was found in all isolates.
Susceptibility testing and detection of resistance genes
The 59 strains were tested for susceptibility to tetracycline and erythromycin using phenotypic and genotypic methods. Fifty-three strains (90%) were resistant to tetracycline (MIC 8-64 mg/L) and 46 (78%) were constitutively resistant to erythromycin (MIC >128 mg/L: n=44, including SsCA-1; MIC 4 mg/L: n=2, including SsUD). All erythromycin-resistant strains were also tetracycline-resistant. The erm(B) gene was the only erythromycin resistance determinant (Figure), found in 44 of 46 erythromycin-resistant strains. Neither erm(A) nor mef(A) were detected in the two erythromycin-resistant (MIC >128 mg/L) erm(B)-negative strains. Tetracycline resistance genes were distributed as follows: tet(O) (n=38), tet(O/W/32/O) (n=8), tet(W) (n=5); tet(M) (n=1), and tet(O)/tet(M) (n=1).
The presence of the mosaic gene was suspected from incongruent findings in PCR experiments, where a 519 bp amplicon was obtained in 38 strains using primers internal to tet(O) (TETO1 and TETO2), and a 2,038 bp amplicon was obtained in 46 strains (of which eight were negative when internal primers were used) using full-length tet(O) primers (TETOFF2 and TETOFR3). In the latter strains the presence of the mosaic gene tet(O/W/32/O) was confirmed by AluI and HinfI restriction analysis and sequencing of PCR products. Sequence analysis (FM164392) revealed that this gene was 99% identical to the tetracycline resistance gene tet(O/W/32/O) (EF065523.1) of an uncultured bacterium isolated from pig faeces [32]. The tet(W) gene was detected in three pig isolates and in both human isolates by HinfI restriction analysis of the amplicons obtained with the tetWFF and tetWFR primer pair and sequencing. Sequence analysis (FN356743) disclosed that it was 99% identical to the tetracycline resistance gene tet(W) (DQ519395.1) of a porcine isolate of Arcanobacterium pyogenes [38].
PFGE typing and MLST
All strains were PFGE-typed after SmaI digestion of total DNA. Thirty-four different pulsotypes were detected and grouped into 14 PFGE types (types 1 to 14) on the basis of a cut-off of 70% similarity (Figure). PFGE type 1 accounted for 52% of isolates and comprised eight pulsotypes (types a to h), of which pulsotype 1a was shared by 22 pig isolates collected from 10 different herds in northern and central Italy in the period from 2003 to 2007. Pulsotype 1d was shared by the human strain SsCA-1 (isolated in 2007) and the pig isolate 10584 (isolated in 2006), and pulsotype 1f was displayed by the human strain SsUD. Comparison of 1d with both pulsotypes 1a and 1f yielded a two-band difference, and comparison of 1a with 1f a three-band difference. MLST of strains v20 (chosen as representative of pulsotype 1a), SsCA-1 (1d), and SsUD (1f) identified the same allelic profile, corresponding to ST1.
Clones
The distribution of cps genes, virulence-associated genes, and tetracycline and erythromycin resistance determinants among the 59 S. suis strains subdivided by PFGE types and pulsotypes is detailed in the Figure. S. suis isolates with a unique combination of a given PFGE pulsotype, a given cps gene, a given virulence profile, and a given resistance genotype and phenotype were considered to represent a clone. According to this criterion, 34 different clones, corresponding to the 34 different pulsotypes, were recognised, 32 of which were found among the 57 pig isolates (Figure). A major cps2 swine clone (clone 1a: mrp, epfclass I; ofstype 1, sly, arcA; tet(O) erm(B)) accounted for 37% of the 59 isolates. Moreover, clones 1d (mrp, epf; ofstype 1, sly, arcA; tet(W) erm(B)) and 1f (mrp, epf; ofstype 1S, sly, arcA; tet(W) erm(B)), containing the two human isolates (SsCA-1 and SsUD, respectively), were seen to be closely related.
Discussion and conclusion
This is the first study of virulence and resistance traits in swine and human strains of S. suis in Italy. The cps genes coding for the capsular polysaccharide as well as mrp, epf, ofs, and sly genes were investigated. The most prevalent capsular gene was cps2, followed by cps9 and cps1. The cps2 and cps9 genes were detected more frequently among invasive isolates; NT isolates were more frequent among non-invasive isolates.
In the present study, virulence-associated genes mrp, epf, sly, and ofs were found in a large proportion of isolates, including NT isolates. The arcA gene was seen in all strains, confirming previous studies [1]. The epf gene was not detected in cps9 strains, in line with a previous report [11], whereas the recently described ofs gene [12,13] was detected not only in all cps2 but also in some cps1, cps9, and NT strains. Human and pig cps2 isolates carrying mrp and epf, were detected. Interestingly, strains carrying mrp and epf have been previously proved to induce meningitis and septicaemia in experimentally infected pigs [39]. Moreover, cps2 strains carrying mrp epfclass I and ofstype 1 were detected in pig isolates. The size variants mrp and epfclass I have been described in human isolates in Europe [40] and recently found in invasive cps2 swine clones from Europe and Brazil [11,41]. The size variant ofstype 1 has been found to be associated with the ST1 complex [13]. Other profiles, such as cps1 mrpS- and cps9 mrp*- have also been described in isolates from diseased pigs in European countries [10,11].
The finding that invasive and non-invasive isolates share identical virulence profiles seems to support the hypothesis that other, as yet unknown virulence factors are involved in S. suis pathogenesis [1,3]. The high allele variability of these genes was confirmed by detection of several size variants of mrp, epf, and ofs, of which some had previously been described [10,11,13,40] and some were new (epf915and ofstype 1S).
High rates of resistance to macrolides and tetracyclines suggested widespread resistance to these antibiotics in Italy. In Europe, rising rates of resistance have been attributed to intensive use by swine breeders of the macrolide-class antibiotic tylosin as a growth promoter and of tetracycline as a therapeutic agent [15]. Co-resistance to macrolides and tetracyclines can be explained by the fact that tetracycline and erythromycin resistance determinants are often linked on mobile genetic elements [42].
All strains were PCR screened for erm(A), erm(B), and mef(A). Neither erm(A) nor mef(A) were detected. The erm(B) gene was found in all but two erythromycin-resistant pig strains, confirming its prevalence in S. suis in Europe [18,19]. A possible explanation for the erythromycin-resistant, erm(A)-, erm(B)- and mef(A)-negative strains could be an erythromycin resistance determinant previously unreported in S. suis [21]. The presence of erm(B) in both human isolates is consistent with its dissemination in the Italian swine population. The genetic basis of erythromycin resistance in human S. suis isolates has barely been investigated [5,21]. The very recent paper by Chu et al. [21] describes the prevalence of mef(A) in isolates from Hong Kong. Interestingly, all mef(A) isolates belonged to ST7 (endemic in Asia) whereas the only erm(B) strain belonged to ST1 (spread worldwide, including in Europe) [21].
The tet(M) and tet(O) genes are common resistance determinants in S. suis, found worldwide both in pig and in human isolates [19,20]. In this study, four tet genes, all coding for ribosomal protection proteins (http://faculty.washington.edu/marilynr/), were found in the Italian S. suis population. While tet(O) was prevalent, tet(M) was, inexplicably, almost absent. In addition tet(W), and the mosaic tet(O/W/32/O), the tet gene found most frequently in pig isolates after tet(O), were detected. The tet(W) gene is associated with tetracycline resistance in a wide range of bacterial species, including obligate anaerobic rumen bacteria and isolates from human gut and oral mucosa. tet(W) was first detected in S. suis by our group in the human isolate SsCA-1 [5], and then here in the other human strain (SSUD) and in some pig isolates. These data suggest that tet(W) could be widespread in S. suis.
The mosaic gene tet(O/W/32/O) has not been described in the genus Streptococcus before. Mosaic tet genes, originating from tet(O) and tet(W), were first detected in 2003 in anaerobic Gram-negative Megasphaera elsdenii from swine intestine [43,44]. Other mosaic genes, also comprising tet(32), were later detected in Clostridium difficile [45]. Initially thought to be confined to a small group of anaerobic bacteria [22], mosaic tet genes have now been found to be abundant in human and animal faecal samples [32] and have also been detected in Bifidobacterium thermophilum and Lactobacillus johnsonii isolates [46]. Further studies on the genetic elements carrying tet genes are warranted to explain the atypical tet distribution observed in Italian S. suis isolates.
Overall, the S. suis pig isolates demonstrated a high genetic diversity that correlates with a wide distribution of S. suis in Italy. In a heterogeneous background population, an identical virulence and resistance profile (cps2 mrp epfclass I ofstype 1 sly erm(B) tet(O)) and pulsotype were shared by more than a third of swine isolates, collected between 2003 and 2007 from different Italian herds and regions, demonstrating the presence and persistence of a dominant clone, 1a.
The results further revealed that the two human isolates shared a number of common or related features, i.e. both were serotype 2 and harboured cps2, both were resistant to erythromycin (MIC 4 μg/ml and >128 μg/ml, respectively) and contained the erm(B) gene, and both were resistant to tetracycline (MIC 16 μg/ml) and contained the tet(W) gene. Moreover, while sharing the same mrp and epf variants as well as sly, the two human isolates SsUD and SsCA-1 bore two different ofs variants, respectively ofstype 1 and ofstype 1S, a new variant with a 324 bp deletion in the ofstype 1 coding sequence.
According to Tenover’s criteria [37], a close relatedness between SsUD and SsCA-1 and between each human isolates and the dominant swine clone was documented by PFGE analysis which yielded pulsotypes with a difference in only two or three bands. MLST analysis assigned clones 1a and 1f (SsUD) to ST1 of the highly virulent ST1 complex, as previously demonstrated also for SsCA-1 (clone 1d) [5]. Overall, our data show that typical Eurasian strains, i.e. strains carrying genes coding for MRP, EF, and suilysin and belonging to the ST1 complex [1], are widespread in Italy.
In conclusion, this study demonstrated a high genetic diversity of Italian S. suis isolates, with a prevalent cps2, erm(B), tet(O) ST1 clone persistent in the swine population. It also demonstrated a close relatedness between two recently isolated cps2 erm(B) and tet(W) ST1 human strains and between human isolates and the dominant swine clone. Finally, it is the first report to demonstrate tet(O/W/32/O) in S. suis and suggests that mosaic tet genes should be sought in S. suis and in other streptococci.
Acknowledgments
This work was partly supported by a grant from the Italian Ministry of Education, University and Research.
References
- Gottschalk M, Segura M, Xu J. Streptococcus suis infections in humans: the Chinese experience and the situation in North America. Anim Health Res Rev. 2007;8(1):29-45.
- Lun ZR, Wang QP, Chen XG, Li AX, Zhu XQ. Streptococcus suis: an emerging zoonotic pathogen. Lancet Infect Dis. 2007;7(3):201-9.
- Wertheim HF, Nghia HD, Taylor W, Schultsz C. Streptococcus suis: an emerging human pathogen. Clin Infect Dis. 2009;48(5):617-25.
- Smith TC, Capuano AW, Boese B, Myers KP, Gray GC. Exposure to Streptococcus suis among US swine workers. Emerg Infect Dis. 2008;14(12):1925-7.
- Manzin A, Palmieri C, Serra C, Saddi B, Princivalli MS, Loi G et al. Streptococcus suis meningitis without evidence of animal contact, Italy. Emerg Infect Dis. 2008;14(12):1946-8.
- King SJ, Leigh JA, Heath PJ, Luque I, Tarradas C, Dowson CG et al. Development of a multilocus sequence typing scheme for the pig pathogen Streptococcus suis: identification of virulent clones and potential capsular serotype exchange. J Clin Microbiol. 2002;40(10):3671-80.
- Ye C, Zhu X, Jing H, Du H, Segura M, Zheng H et al. Streptococcus suis sequence type 7 outbreak, Sichuan, China. Emerg Infect Dis. 2006;12(8):1203-8.
- Staats JJ, Feder I, Okwumabua O, Chengappa MM. Streptococcus suis: past and present. Vet Res Commun. 1997;21(6):381-407.
- Hill JE, Gottschalk M, Brousseau R, Harel J, Hemmingsen SM, Goh SH. Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet Microbiol. 2005;107(1-2):63-9.
- Wisselink HJ, Smith HE, Stockhofe-Zurwieden N, Peperkamp K, Vecht U. Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Vet Microbiol. 2000;74(3):237-48.
- Silva LM, Baums CG, Rehm T, Wisselink HJ, Goethe R, Valentin-Weigand P. Virulence-associated gene profiling of Streptococcus suis isolates by PCR. Vet Microbiol. 2006;115(1-3):117-27.
- Baums CG, Kaim U, Fulde M, Ramachandran G, Goethe R, Valentin-Weigand P. Identification of a novel virulence determinant with serum opacification activity in Streptococcus suis. Infect Immun. 2006;74(11):6154-62.
- Takamatsu D, Osaki M, Tharavichitkul P, Takai S, Sekizaki T. Allelic variation and prevalence of serum opacity factor among the Streptococcus suis population. J Med Microbiol. 2008;57(Pt4):488-94.
- Gruening P, Fulde M, Valentin-Weigand P, Goethe R. Structure, regulation, and putative function of the arginine deiminase system of Streptococcus suis. J Bacteriol. 2006;188(2):361-9
- Wisselink HJ, Veldman KT, Van den Eede C, Salmon SA, Mevius DJ. Quantitative susceptibility of Streptococcus suis strains isolated from diseased pigs in seven European countries to antimicrobial agents licensed in veterinary medicine. Vet Microbiol. 2006;113(1-2):73-82
- Hendriksen RS, Mevius DJ, Schroeter A, Teale C, Jouy E, Butaye P et al. Occurrence of antimicrobial resistance among bacterial pathogens and indicator bacteria in pigs in different European countries from year 2002 – 2004: the ARBAO-II study. Acta Vet Scand. 2008;13(1);50:19.
- Zhang C, Ning Y, Zhang Z, Song L, Qiu H, Gao H. In vitro antimicrobial susceptibility of Streptococcus suis strains isolated from clinically healthy sows in China. Vet Microbiol. 2008;131(3-4):386-92.
- Martel A, Baele M, Devriese LA, Goossens H, Wisselink HJ, Decostere A, et al. Prevalence and mechanism of resistance against macrolides and lincosamides in Streptococcus suis isolates. Vet Microbiol. 2001;83(3):287-97.
- Tian Y, Aarestrup FM, Lu CP. Characterization of Streptococcus suis serotype 7 isolates from diseased pigs in Denmark. Vet Microbiol. 2004;103(1-2):55-62.
- Ye C, Bai X, Zhang J, Jing H, Zheng H, Du H, et al. Spread of Streptococcus suis sequence type 7, China. Emerg Infect Dis 2008;14(5):787-91
- Chu YW, Cheung TK, Chu MY, Tsang VY, Fung JT, Kam KM, et al. Resistance to tetracycline, erythromycin and clindamycin in Streptococcus suis serotype 2 in Hong Kong. Int J Antimicrob Agents. 2009;34(2):181-2.
- Roberts MC. Update on acquired tetracycline resistance genes. FEMS Microbiol Lett. 2005;245(2):195-203.
- Perseghin P, Bezzi G, Troupioti P, Gallina M. Streptococcus suis meningitis in an Italian blood donor. Lancet. 1995;346(8985):1305-6.
- Camporese A, Tizianel G, Bruschetta G, Cruciatti B, Pomes A. Human meningitis caused by Streptococcus suis: the first case report from north-eastern Italy. Infez Med. 2007;15(2):111-4 [Article in Italian].
- Clsi.org [homepage on the internet]. Wayne, PA. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing. 18th informational supplement. Matthew A Wikler et al. Clinical and Laboratory Standards Institute; 2008. CLSI Document M100-S18.
- Clsi.org [homepage on the internet]. Wayne, PA. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial disk and dilution susceptibility tests for bacteria isolated from animals, approved standard, 2nd Edition. Clinical and Laboratory Standards Institute; 2002. CLSI Document M31-A2. Available from: http://www.clsi.org/source/orders/free/m31-a3.pdf
- Giovanetti E, Montanari MP, Mingoia M, Varaldo PE. Phenotypes and genotypes of erythromycin-resistant Streptococcus pyogenes strains in Italy and heterogeneity of inducibly resistant strains. Antimicrob Agents Chemother. 1999;43(8):1935-40.
- Sutcliffe J, Tait-Kamradt A, Wondrack L. Streptococcus pneumoniae and Streptococcus pyogenes resistant to macrolides but sensitive to clindamycin: a common resistance pattern mediated by an efflux system. Antimicrob Agents Chemother. 1996;40(8):1817-24.
- Seppälä H, Skurnik M, Soini H, Roberts MC, Huovinen P. A novel erythromycin resistance methylase gene (ermTR) in Streptococcus pyogenes. Antimicrob Agents Chemother. 1998;42(2):257-62.
- Trzcinski K, Cooper BS, Hryniewicz W, Dowson CG. Expression of resistance to tetracyclines in strains of methicillin-resistant Staphylococcus aureus. J Antimicrob Chemother. 2000;45(6):763-70.
- Olsvik B, Olsen I, Tenover FC. Detection of tet(M) and tet(O) using the polymerase chain reaction in bacteria isolated from patients with periodontal disease. Oral Microbiol Immunol. 1995;10(4):87-92.
- Patterson AJ, Rincon MT, Flint HJ, Scott KP. Mosaic tetracycline resistance genes are widespread in human and animal fecal samples. Antimicrob Agents Chemother. 2007;51(3):1115-8.
- Aminov RI, Garrigues-Jeanjean N, Mackie RI. Molecular ecology of tetracycline resistance: development and validation of primers for detection of tetracycline resistance genes encoding ribosomal protection proteins. Appl Environ Microbiol. 2001;67(1):22-32.
- Vela AI, Goyache J, Tarradas C, Luque I, Mateos A, Moreno MA et al. Analysis of genetic diversity of Streptococcus suis clinical isolates from pigs in Spain by pulsed-field gel electrophoresis. J Clin Microbiol. 2003;41(6):2498-502.
- Berthelot-Hérault F, Marois C, Gottschalk M, Kobisch M. Genetic diversity of Streptococcus suis strains isolated from pigs and humans as revealed by pulsed-field gel electrophoresis. J Clin Microbiol. 2002;40(2):615-9.
- Allgaier A, Goethe R, Wisselink HJ, Smith HE, Valentin-Weigand P. Relatedness of Streptococcus suis isolates of various serotypes and clinical backgrounds as evaluated by macrorestriction analysis and expression of potential virulence traits. J Clin Microbiol. 2001;39(2):445-53.
- Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH et al. Interpreting chromosomal DNA restriction patterns. J Clin Microbiol. 1995;33(9):2233-9.
- Billington SJ, Jost BH. Multiple genetic elements carry the tetracycline resistance gene tet(W) in the animal pathogen Arcanobacterium pyogenes. Antimicrob Agents Chemother. 2006;50(11):3580-7
- Smith HE, Vecht U, Wisselink HJ, Stockhofe-Zurwieden N, Biermann Y, Smits MA. Mutants of Streptococcus suis types 1 and 2 impaired in expression of muramidase-released protein and extracellular protein induce disease in newborn germfree pigs. Infect Immun. 1996;64(10):4409-12.
- Smith HE, Reek FH, Vecht U, Gielkens AL, Smits MA. Repeats in an extracellular protein of weakly pathogenic strains of Streptococcus suis type 2 are absent in pathogenic strains. Infect Immun. 1993;61(8):3318-26.
- Martinez G, Pestana de Castro AF, Ribeiro Pagnani KJ, Nakazato G, Dias da Silveira W, Gottschalk M. Clonal distribution of an atypical MRP+, EF*, and suilysin+ phenotype of virulent Streptococcus suis serotype 2 strains in Brazil. Can J Vet Res. 2003;67(1):52-5
- Roberts MC. Update on macrolide-lincosamide-streptogramin, ketolide, and oxazolidinone resistance genes. FEMS Microbiol Lett. 2008;282(2):147-59.
- Stanton TB, Humphrey SB. Isolation of tetracycline-resistant Megasphaera elsdenii strains with novel mosaic gene combinations of tet(O) and tet(W) from swine. Appl Environ Microbiol. 2003;69(7):3874-82.
- Stanton TB, McDowall JS, Rasmussen MA. Diverse tetracycline-resistant genotypes of Megasphaera elsdenii strains selectively cultured from swine feces. Appl Environ Microbiol. 2004;70(6):3754-7.
- Spigaglia P, Barbanti F, Mastrantonio P. Tetracycline resistance gene tet(W) in the pathogenic bacterium Clostridium difficile. Antimicrob Agents Chemother. 2008;52(2):770-3.
- van Hoek AH, Mayrhofer S, Domig KJ, Flórez AB, Ammor MS, Mayo B et al. Mosaic tetracycline resistance genes and their flanking regions in Bifidobacterium thermophilum and Lactobacillus johnsonii. Antimicrob Agents Chemother. 2008;52(1):248-52.
Outbreak associated with Streptococcus suis in pigs in China
3 August 2005
To date, the Ministry of Health in China has reported 206 cases of human
disease associated with an outbreak of Streptococcus suis in pigs. Of
these human cases, 38 have been fatal. As reported by China, 18 patients
are critically ill.
Virtually all cases have occurred in Sichuan Province, where infections
with Streptococcus suis have been detected in pigs in a concurrent
outbreak. The province has one of the largest pig populations in China.
Investigation and containment of the outbreak have been given high
priority by Chinese authorities. The country’s ministries of health and
agriculture are working in close collaboration, and WHO and FAO are
being promptly informed of new developments.
Investigations conducted by Chinese epidemiologists indicate that the
first human cases occurred at the end of June in Ziyang City, Sichuan
Province. From 24 June through 21 July, the authorities reported 20
cases of illness, of unknown cause, admitted to three hospitals in that
city. WHO was officially informed of the outbreak on 22 July, at which
time 20 cases and 9 deaths had been reported.
Cases have since been reported in 11 prefectures in Sichuan Province.
Most cases reported have occurred in adult male farmers. Information
reported to WHO suggests that close contact with diseased or dead pigs
is the principal source of human infection.
Symptoms reported by local clinicians include high fever, malaise,
nausea, and vomiting, followed by meningitis, subcutaneous haemorrhage,
toxic shock, and coma in severe cases. The incubation period is short
and disease progression is rapid.
Local experts are conducing active searches for further cases. To date,
Chinese authorities say they have found no evidence of human-to-human
transmission.
The outbreak in humans has some unusual features and is being closely
followed by WHO. Diagnostic testing to further characterize the
causative agent is recommended as an essential part of ongoing efforts
to understand this outbreak, ensure its rapid containment, and prevent
further deaths.